Where on the Phylogenetic Tree Should the Origin of Alveoli Structure Appear?

A highly resolved, mechanically generated tree of life, settled along all sequenced genomes.[1] [2]
A phylogenetic tree (also phylogeny or evolutionary tree [3]) is a branching diagram or a tree showing the biological process relationships among various biological species operating theatre other entities based upon similarities and differences in their physical operating theatre genetic characteristics. All life on Earth is part of a single phylogenetic corner, indicating shared ancestry.
In a stock-still phylogenetic tree, each thickening with descendants represents the inferred most recent common antecedent of those descendants,[ citation necessary ] and the edge lengths in some trees may be interpreted As time estimates. Each node is called a assortment unit. Internal nodes are generally called suppositional taxonomic units, every bit they cannot be straight off ascertained. Trees are utilitarian in Fields of biology such atomic number 3 bioinformatics, systematics, and phylogenetics. Unrooted trees instance only the relatedness of the riffle nodes and do not require the ancestral solution to be known or inferred.
Story [edit]
The idea of a "shoetree of animation" arose from ancient notions of a ladder-look-alike progression from depress into higher forms of life (such as in the Great Chain of Being). Early representations of "fork" biological process trees include a "earth science chart" screening the geological relationships among plants and animals in the Word Primary Geology, by Edward Hitchcock (first variant: 1840).
Charles Charles Robert Darwin (1859) also produced one of the best illustrations and crucially popularized the notion of an evolutionary "tree" in his bodily fluid book The Origin of Species. Ended a century later, evolutionary biologists however use tree diagrams to depict phylogenesis because so much diagrams effectively convey the concept that speciation occurs through the adaptive and semirandom splitting of lineages. Complete time, species classification has become less static and to a greater extent dynamic.
The term phylogenetic, or phylogeny, derives from the two ancient Grecian words φῦλον ( phûlon ), meaningful "belt along, lineage", and γένεσις ( génesis ), meaning "root, source".[4] [5]
Properties [edit]
Rooted tree [blue-pencil]

Rooted phylogenetic tree optimized for blind people. The worst point of the tree is the root, which symbolizes the cosmopolitan common antecedent to all living beings. The tree diagram branches out into troika important groups: Bacteria (left ramification, letters a to i), Archea (middle fork, letters j to p) and Eukaryota (right branch, letters q to z). Each varsity letter corresponds to a group of organisms, traded below this description. These letters and the description should be converted to Braille font, and printed using a Braille printer. The project can buoy be 3D printed away copying the png file and using Cura or other software to generate the Gcode for 3D printing process.
A rooted phylogenetic Sir Herbert Beerbohm Tree (see cardinal graphics at top) is a directed tree with a unique node — the root — corresponding to the (usually imputed) almost Recent epoch common ascendent of all the entities at the leaves of the Tree. The root node does not have a parent node, but serves as the parent of all other nodes in the tree. The origin is therefore a node of degree 2, while another interior nodes have a borderline academic degree of 3 (where "degree" Hera refers to the total routine of incoming and outbound edges).
The most commons method acting for rooting trees is the use of an uncontroversial outgroup—close enough to allow illation from trait data or molecular sequencing, but far plenty to be a clear outgroup.
Unrooted Tree [delete]

Unrooted trees illustrate the relatedness of the leaf nodes without making assumptions about ancestry. They do not require the hereditary root to be known or inferred.[7] Unrooted trees can e'er constitute generated from rooted ones by merely omitting the root. By contrast, inferring the root of an unrooted tree requires some means of distinguishing ancestry. This is normally done by including an outgroup in the input data and so that the root is necessarily between the outgroup and the rest of the taxa in the tree, or by introducing additional assumptions about the relative rates of evolution on each subdivision, such as an coating of the unit time hypothesis.[8]
Bifurcating versus multifurcating [edit]
Both rooted and unrooted trees can be either bifurcating or multifurcating. A rooted bifurcating tree has on the nose cardinal descendants arising from each indoor node (that is, it forms a multiple shoetree), and an unrooted bifurcating shoetree takes the form of an unrooted binary tree, a free tree with exactly three neighbors at each internal node. In contrast, a rooted multifurcating tree diagram may have more than cardinal children at some nodes and an unrooted multifurcating tree may get more than three neighbors at some nodes.
Labeled versus unlabelled [blue-pencil]
Both rooted and unrooted trees john be either tagged OR unlabeled. A labeled tree has specific values assigned to its leaves, while an unlabeled tree, sometimes called a tree shape, defines a topology only. Or s succession-based trees built from a small genomic locus, such as Phylotree,[9] feature interior nodes labelled with inferred heritable haplotypes.
Enumerating trees [edit]

Increase in the total number of phylogenetic trees as a occasion of the number of labeled leaves: unrooted positional representation system trees (blue diamonds), rooted binary trees (red circles), and rooted multifurcating or binary trees (common: triangles). The Y-axis ordered series is logarithmic.
The number of attainable trees for a given number of leaf nodes depends happening the specific typewrite of tree, but there are ever more labeled than unlabeled trees, more multifurcating than bifurcating trees, and more nonmoving than unrooted trees. The last distinction is the almost biologically relevant; it arises because there are numerous places connected an unrooted tree to put the root. For bifurcating labeled trees, the total number of rooted trees is:
- for , represents the number of leaf nodes.[10]



For bifurcating tagged trees, the total number of unrooted trees is:[10]
- for .


Among tagged bifurcating trees, the number of unrooted trees with leaves is equal to the number of rooted trees with leaves.[3]

The keep down of vegetable trees grows quickly as a function of the number of tips. For 10 tips, thither are to a higher degree likely bifurcating trees, and the number of multifurcating trees rises quicker, with ca. 7 times as many of the latter atomic number 3 of the former.

| Tagged leaves | Binary unrooted trees | Binary rooted trees | Multifurcating rooted trees | All possible rooted trees |
|---|---|---|---|---|
| 1 | 1 | 1 | 0 | 1 |
| 2 | 1 | 1 | 0 | 1 |
| 3 | 1 | 3 | 1 | 4 |
| 4 | 3 | 15 | 11 | 26 |
| 5 | 15 | 105 | 131 | 236 |
| 6 | 105 | 945 | 1,807 | 2,752 |
| 7 | 945 | 10,395 | 28,813 | 39,208 |
| 8 | 10,395 | 135,135 | 524,897 | 660,032 |
| 9 | 135,135 | 2,027,025 | 10,791,887 | 12,818,912 |
| 10 | 2,027,025 | 34,459,425 | 247,678,399 | 282,137,824 |
Unscheduled tree types [edit]

Dendrogram of the evolution of some blackguard breeds
Dendrogram [cut]
A dendrogram is a systemic appoint for a Sir Herbert Beerbohm Tree, whether organic process or not, and thence also for the diagrammatic representation of a phylogenetic tree.[11]
Cladogram [edit]
A cladogram only represents a branching pattern; i.e., its offset lengths do not represent time or relative amount of fibre deepen, and its internecine nodes do not represent ancestors.[12]

A chronogram of Lepidoptera.[13] In this phylogenetic tree diagram type, offshoot lengths are proportional to geologic time.
Phylogram [edit]
A phylogram is a biological process tree diagram that has branch lengths proportional to the amount of lineament change.[14]
A chronogram is a phylogenetic tree that explicitly represents time through its offset lengths.[15]
Dahlgrenogram [edit]
A Dahlgrenogram is a diagram representing a cross part of a phylogenetic tree
Phyletic network [blue-pencil]
A phylogenetic web is not strictly speaking a tree, but rather a Thomas More general graph, or a directed acyclic graph in the case of rooted networks. They are utilised to overcome some of the limitations inherent to trees.
Mandril diagram [edit]
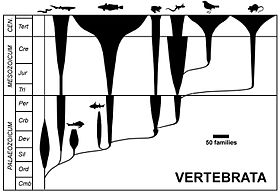
A arbo diagram, showing the evolution of the vertebrates at class level, width of spindles indicating number of families. Spindle diagrams are often used in biological process taxonomy.
A spindle diagram, or bubble diagram, is often called a romerogram, after its popularisation by the American palaeontologist Alfred Romer.[16] It represents taxonomic diversity (horizontal width) against earth science time (plumb axis) in order to reflect the variation of teemingness of individual taxa direct time. However, a spindle diagram is not an organic process tree:[17] the taxonomic spindles obscure the actual relationships of the parent taxon to the girl taxon[16] and have the disadvantage of involving the paraphyly of the paternal group.[18] This case of diagram is no longer victimized in the form in the beginning proposed.[18]
Chromatic of life [edit]

Darwin[19] also mentioned that the coral Crataegus laevigata be a more suitable metaphor than the tree. Indeed, phylogenetic corals are useful for portraying bygone and on hand life, and they have some advantages over trees (anastomoses allowed, etc.).[18]
Grammatical construction [edit]
Phylogenetic trees composed with a nontrivial number of input sequences are constructed exploitation computational phylogenetics methods. Distance-matrix methods such as neighbour-joining or UPGMA, which estimate genetic distance from multiple sequence alignments, are simplest to implement, but make not invoke an evolutionary sit. Many sequence alignment methods such as ClustalW also create trees by using the simpler algorithms (i.e. those supported distance) of tree construction. Maximum parsimony is another simple method of estimating phylogenetic trees, but implies an implicit good example of evolution (i.e. parsimony). More late methods use the optimality criterion of maximum likelihood, often within a Bayesian framework, and apply an explicit mold of evolution to phylogenetic tree estimation.[3] Identifying the optimal tree diagram using many of these techniques is NP-granitic,[3] so heuristic search and optimisation methods are used in compounding with tree-marking functions to identify a sensibly good tree that fits the information.
Tree-edifice methods can be assessed on the basis of several criteria:[20]
- efficiency (how long does it take to compute the answer, how often memory does it need?)
- power (does it make good usance of the information, or is entropy being wasted?)
- consistency (will it converge happening the same answer repeatedly, if each time given different information for the same model job?)
- robustness (does IT cope easily with violations of the assumptions of the underlying framework?)
- falsifiability (does IT alert us when it is not good to use, i.e. when assumptions are profaned?)
Tree-building techniques have too gained the attention of mathematicians. Trees can besides be built using T-theory.[21]
File formats [edit]
Trees can be encoded in a figure of different formats, all of which must represent the nested structure of a tree. They may or may non encode branch lengths and other features. Exchangeable formats are critical for distributing and sharing trees without relying on graphics output that is hard to import into existing software. Commonly used formats are
- Nexus file format
- Newick data format
Limitations of phylogenetic analysis [redact]
Although phylogenetic trees produced connected the fundament of sequenced genes or genomic data in different species buns provide evolutionary insight, these analyses have important limitations. Most importantly, the trees that they generate are not necessarily correct – they do not of necessity accurately represent the evolutionary history of the enclosed taxa. As with any scientific result, they are subject to falsification by further study (e.g., gathering of additional data, analyzing the existing data with improved methods). The data happening which they are based may be noisy;[22] the depth psychology can be confounded aside genetic recombination,[23] horizontal gene transfer,[24] hybridisation between species that were non nearest neighbors on the tree earlier hybridisation takes place, convergent evolution, and preserved sequences.
Also, there are problems in basing an analysis on a single type of character, so much as a single gene or protein or just on morphological analytic thinking, because so much trees constructed from another unrelated data source ofttimes differ from the first, and therefore great care is required in inferring phylogenetic relationships among species. This is almost true of genetic material that is subject to lateral gene transfer and recombination, where different haplotype blocks can have different histories. In these types of analytic thinking, the output tree of a phylogenetic psychoanalysis of a single cistron is an estimate of the gene's evolution (i.e. a gene tree) and not the phylogeny of the taxa (i.e. species tree) from which these characters were sampled, though ideally, both should be rattling close-knit. For this reason, serious phylogenetic studies generally use a compounding of genes that come from different genomic sources (e.g., from mitochondrial or plastid vs. nuclear genomes),[25] or genes that would personify expected to develop low-level different exclusive regimes, so that homoplasy (false homology) would be unlikely to result from natural choice.
When extinct species are enclosed every bit terminal nodes in an analysis (rather than, for example, to constrain internal nodes), they are considered non to represent direct ancestors of whatever extant species. Extinct species do not typically contain mellow-upper-class Desoxyribonucleic acid.
The range of useful DNA materials has distended with advances in extraction and sequencing technologies. Development of technologies able to deduce sequences from smaller fragments, OR from spatial patterns of Deoxyribonucleic acid degradation products, would further expand the range of Deoxyribonucleic acid considered useful.
Phyletic trees can also embody inferred from a stove of other data types, including morphology, the presence or absence of especial types of genes, insertion and deletion events – and any other observation thought to contain an evolutionary signal.
Phylogenetic networks are used when bifurcating trees are not suitable, imputable these complications which propose a Thomas More reticulate evolutionary history of the organisms sampled.
See too [edit]
- Clade
- Cladistics
- Computational phylogenetics
- Biological process biology
- Biological process taxonomy
- Generalized Sir Herbert Beerbohm Tree alignment
- List of phylogenetics computer software
- List of phylogenetic tree visualization software
- PANDIT, a biological database screening protein domains
- Phylogenetic comparative methods
References [delete]
- ^ Letunic, Ivica; Bork, Peer (1 January 2007). "Interactive Tree Of Life (iTOL): an online tool for phylogenetic tree display and annotation" (PDF). Bioinformatics. 23 (1): 127–128. Interior Department:10.1093/bioinformatics/btl529. ISSN 1367-4803. PMID 17050570. Archived (PDF) from the original on November 29, 2015. Retrieved 2015-07-21 .
- ^ Ciccarelli, F. D.; Doerks, T.; Von Mering, C.; Creevey, C. J.; Snel, B.; Bork, P. (2006). "Toward automatic Reconstruction Period of a extremely resolved tree of liveliness" (PDF). Science. 311 (5765): 1283–1287. Bibcode:2006Sci...311.1283C. CiteSeerX10.1.1.381.9514. doi:10.1126/science.1123061. PMID 16513982. S2CID 1615592.
- ^ a b c d Felsenstein J. (2004). Inferring Phylogenies Sinauer Associates: Sunderland, MA.
- ^ Bailly, Anatole (1981-01-01). Abrégé du dictionnaire grec français. Paris: Hachette. ISBN978-2010035289. OCLC 461974285.
- ^ Bailly, Anatole. "Greek-European country dictionary online". www.tabularium.be. Archived from the freehanded on April 21, 2014. Retrieved Master of Architecture 2, 2018.
- ^ Hodge T, Cope M (1 October 2000). "A myosin family tree". J Cell Sci. 113 (19): 3353–4. Interior Department:10.1242/jcs.113.19.3353. PMID 10984423. Archived from the original on 30 September 2007.
- ^ ""Tree" Facts: Rooted versus Unrooted Trees". Archived from the original on 2014-04-14. Retrieved 2014-05-26 .
- ^ Maher BA (2002). "Uprooting the Tree of Life". The Scientist. 16 (2): 90–95. Bibcode:2000SciAm.282b..90D. DoI:10.1038/scientificamerican0200-90. PMID 10710791. Archived from the original on 2003-10-02.
- ^ van Oven, Mannis; Kayser, Manfred (2009). "Updated spatiotemporal phylogenetic tree diagram of global human mitochondrial DNA variation". Humanlike Genetic mutation. 30 (2): E386–E394. doi:10.1002/humu.20921. PMID 18853457. S2CID 27566749.
- ^ a b c Felsenstein, Joseph (1978-03-01). "The Number of Evolutionary Trees". Systematic Biology. 27 (1): 27–33. doi:10.2307/2412810. ISSN 1063-5157. JSTOR 2412810.
- ^ Fox, Emily. "The dendrogram". coursea. coursea. Archived from the original on 28 September 2017. Retrieved 28 September 2017.
- ^ Mayr, Ernst (2009)"Cladistics or cladistic classification?". Daybook of Zoological Systematics and Evolutionary Research. 12: 94–128. doi:10.1111/j.1439-0469.1974.tb00160.x..
- ^ Labandeira, C. C.; Dilcher, D. L.; Davis, D. R.; Wagner, D. L. (1994-12-06). "97 jillio years of angiosperm-insect association: paleobiological insights into the meaning of coevolution". Proceedings of the National Academy of Sciences. 91 (25): 12278–12282. Bibcode:1994PNAS...9112278L. Department of the Interior:10.1073/pnas.91.25.12278. ISSN 0027-8424. PMC45420. PMID 11607501.
- ^ Soares, Antonio; Râbelo, Ricardo; Delbem, Alexandre (2017). "Optimization based on phylogram analysis". Practiced Systems with Applications. 78: 32–50. doi:10.1016/j.eswa.2017.02.012. ISSN 0957-4174.
- ^ Santamaria, R.; Theron, R. (2009-05-26). "Treevolution: visual analysis of phylogenetic trees". Bioinformatics. 25 (15): 1970–1971. doi:10.1093/bioinformatics/btp333. PMID 19470585.
- ^ a b "Organic process systematics: Arbo Diagrams". Palaeos.com. 2014-11-10. Retrieved 2019-11-07 .
- ^ "Trees, Bubbles, and Hooves". A Three-Pound Monkey around Brain — Biology, programming, philology, organic evolution, systematics …. 2007-11-21. Retrieved 2019-11-07 .
- ^ a b c Podani, János (2019-06-01). "The Coral of Life". Evolutionary Biology. 46 (2): 123–144. doi:10.1007/s11692-019-09474-w. ISSN 1934-2845.
- ^ Charles Robert Darwin, Charles (1837). Notebook B. p. 25.
- ^ Penny, D.; Hendy, M. D.; Steel, M. A. (1992). "Progress with methods for constructing evolutionary trees". Trends in Ecology and Phylogenesis. 7 (3): 73–79. doi:10.1016/0169-5347(92)90244-6. PMID 21235960.
- ^ A. Dress, K. T. Huber, and V. Moulton. 2001. Metric Spaces in Pure and Applied Mathematics. Documenta Mathematica LSU 2001: 121-139
- ^ Townsend JP, Su Z, Tekle Y (2012). "Phylogenetic Signal and Make noise: Predicting the Exponent of a Information Set to Firmness of purpose Phylogeny". Genetics. 61 (5): 835–849. doi:10.1093/sysbio/sys036. PMID 22389443.
- ^ Arenas M, Posada D (2010). "The effect of recombination on the reconstructive memory of patrimonial sequences". Genetics. 184 (4): 1133–1139. doi:10.1534/genetics.109.113423. PMC2865913. PMID 20124027.
- ^ Woese C (2002). "On the evolution of cells". Proc Natl Acad Sci USA. 99 (13): 8742–7. Bibcode:2002PNAS...99.8742W. doi:10.1073/pnas.132266999. PMC124369. PMID 12077305.
- ^ Parhi, J.; Tripathy, P.S.; Priyadarshi, H.; Mandal, S.C.; Pandey, P.K. (2019). "Diagnosis of mitogenome for robust phylogeny: A case of Cypriniformes fish group". Gene. 713: 143967. Department of the Interior:10.1016/j.gene.2019.143967. PMID 31279710.
Encourage recitation [edit]
- Schuh, R. T. and A. V. Z. Brower. 2009. Biological Systematics: principles and applications (2nd edn.) ISBN 978-0-8014-4799-0
- Manuel Lima, The Book of Trees: Visualizing Branches of Knowledge, 2014, Princeton Architectural Press, Fresh York.
- MEGA, a free software program to draw phylogenetic trees.
- Gontier, N. 2011. "Portraying the Tree of Life: the Philosophical and Historical Roots of Organic process Tree Diagrams." Evolution, Teaching, Outreach 4: 515–538.
External golf links [edit]
Images [delete]
- Human Y-Chromosome 2002 Phylogenetic Tree
- iTOL: Interactive Tree Of Living
- Phylogenetic Tree of Artificial Organisms Evolved connected Computers
- Miyamoto and Goodman's Phylogram of Eutherian Mammals
General [edit]
- An overview of different methods of Tree visualisation is getable at Page, R. D. M. (2011). "Space, time, form: Viewing the Tree of Living". Trends in Ecology & Phylogenesis. 27 (2): 113–120. doi:10.1016/j.tree.2011.12.002. PMID 22209094.
- OneZoom: Tree of Life – every living species as nonrational and zoomable fractal IE (responsive design)
- Discover Life An interactive tree supported on the U.S. Status Science Origination's Assembling the Tree of Life Project
- PhyloCode
- A Multiple Alignment of 139 Myosin Sequences and a Phylogenetic Tree
- Tree of Life Web Project
- Phylogenetic inferring on the T-King server
- NCBI's Taxonomy Database[1]
- ETE: A Python Surroundings for Tree Exploration This is a programming library to analyze, manipulate and visualize phylogenetic trees. Ref.
- A daily-updated tree diagram of (sequenced) aliveness Fang, H.; Oates, M. E.; Pethica, R. B.; Greenwood, J. M.; Sardar, A. J.; Rackham, O. J. L.; Donoghue, P. C. J.; Stamatakis, A.; First State Lima Morais, D. A.; Gough, J. (2013). "A daily-updated tree of (sequenced) life sentence as a reference work for genome explore". Knowledge domain Reports. 3: 2015. Bibcode:2013NatSR...3E2015F. doi:10.1038/srep02015. PMC6504836. PMID 23778980.
Where on the Phylogenetic Tree Should the Origin of Alveoli Structure Appear?
Source: https://en.wikipedia.org/wiki/Phylogenetic_tree
0 Response to "Where on the Phylogenetic Tree Should the Origin of Alveoli Structure Appear?"
Postar um comentário